
精選版 日本国語大辞典 「視覚」の意味・読み・例文・類語
し‐かく【視覚】
〘名〙 (gezigt の訳語) 目を受容器として生ずる感覚の総称。光が網膜上にある感覚細胞を刺激して生じる。明暗覚、色覚、形態覚、運動覚などが含まれる。視感。〔和蘭字彙(1855‐58)〕
[語誌]「和蘭字彙」にあるのが古く、蘭学者による造語と考えられる。その後、「英和対訳袖珍辞書」(一八六二)にも複合語を含め、少なからず訳語として使用されており、明治初期には一般化したと思われる。
出典 精選版 日本国語大辞典精選版 日本国語大辞典について 情報
最新 心理学事典 「視覚」の解説
しかく
視覚
vision (英・仏),Gesichtssinn (独)
こうして成立した視覚世界があまりに豊かに感じられるので,網膜像そのものを眺めているかのように感じられがちだが,以下の理由から考えて,脳内で視覚情報の解析が行なわれた結果が,われわれの視覚体験の本質であると結論される。第1に,網膜像として得られる視覚情報は眼の動きに伴いつねに揺れ動き,視野中心に対応する中心窩fovea付近以外は解像度が低く,視野の盲点に対応する視神経円板optic diskや網膜血管系の影の部分においては情報が欠損しているにもかかわらず,われわれの意識する世界は安定的で連続的にくっきりと感じられる。第2に,網膜像を直に意識化しているなら生じないはずの神経心理学的症状として,眼に異常がなくても対象が歪む,二重に見える,距離感がない,など特定の視機能だけが損なわれることがある。第3に,もし心内で網膜像がそのまま映っているなら,いったいだれがそれを解析するのかという哲学的問題があり,心内の映像を心の中の小人的存在であるホムンクルスhomunculusが見て解析すると考えると,そのホムンクルスの心内で同じ問題が生じ,無限後退に陥ってしまう。
視覚世界がいかに構築されるかに関しては諸説あり,解明が待たれる。「世界に関する知識」および「現在までに得られている網膜像の入力データ」から考えられる最も妥当な世界のあり方に関して,ある種の信念belief あるいは「外界に関する予測」が形作られ,時々刻々入力される新しいデータと突き合わせてつねに検証され,予測誤差prediction errorが生じれば新しい信念へと更新される。フリストンFriston,K.はじめ神経科学者や認識哲学者が提唱するこのような大枠のもとで,いかなる形式のモデルが脳内のどこに表象され,われわれの視覚的意識にいかに対応するのかということは,意識の神経相関neural correlate of consciousness(NCC)を探究するうえで大きな問題である。
【不良設定問題と自然制約条件】 網膜像に含まれる情報は,外界構造を計算するのにまったく不十分である。典型例を挙げれば,網膜像は2次元投影像であるため奥行き次元が失われており,同じ網膜像をもたらしうる3次元外界構造には無限の可能性がある。たとえば正方形の網膜像があったとき,その原因となる外界の物体は前額平行面上の正方形かもしれないし,奥行き方向に傾いた長方形かもしれないし,あるいは直方体や四角錐が特定の角度に置かれているのかもしれない。また,ある物体からの表面反射光が得られた場合,その原因となる外界の照明光の分光分布,光量と物体の分光反射率と入射角度の関係は特定できない。ある光強度の物体反射光は,暗い照明が白い表面に当たった結果かもしれないし,明るい照明が薄暗い表面に当たった結果かもしれないし,明るい照明が白い表面に対して斜めに当たったために反射光量が少ないのかもしれない。与えられた情報だけでは解が一意に定まらない計算問題を不良設定問題ill-posed problemといい,最適な解を推定するという作業が必要になる。視覚情報処理系はつねに不良設定問題にさらされ,外界に関する仮定である自然制約条件natural constraintをさまざまにおいて解を絞っている。その重要な2例として,事物の性質は簡単に変わらないはずだとする知覚の恒常性perceptual constancyと,自然画像の統計量は特定の分布に従うはずだとする事前確率prior probabilityが挙げられる。
【視覚の下位区分】 視覚という感覚モダリティは膨大な情報処理を含む大きな概念なので,細分化して理解することが重要な場合がある。異なる感覚属性は独立した装置で処理されるという機能的モジュール性が,フォーダーFodor,J.はじめ多くの研究者によって提唱されたこともあり,明るさ・色・肌理・運動・奥行きなどの感覚属性で区分することが理解の助けになって,各々独立に研究が進んできた。また,空間視spatial visionと形態視form visionの区分や,行為actionと認識recognitionの区分など,異なる処理経路を介すると考えられる計算内容によっても分けられる。
このような並列的区分とは別に,研究分野の違いによる階層的区分も可能である。まず網膜に結像するまでを主に記述する眼光学ophthalmological opticsの分野があり,次にフィルターや並列結合素子などの工学的概念を多用して視覚信号の生体情報表現を扱う初期視覚low-level visionの分野がある。これらに対して,能動的注意,物体認識,シーン解析,視覚的記憶などの高次機能を扱う分野があり,これを後期視覚high-level visionとよんで区別することがある。したがって,ひとくちに空間や時間などの諸特性を記述するうえでも,扱う属性や階層によって大きく特性が異なるので統一的な説明は難しい。
【空間特性】 ヒトやサルなど霊長類では,網膜に中心窩があり視野中心が最も解像度が高く,視野周辺に行くにつれて視力が悪くなる。視力の評価の一つである縞視力grating acuityは,解像しうる最も細かい縞をもって指標とする。縞の細かさは視角1°当たり何周期の輝度変調があるかをもって記述し,これを空間周波数spatial frequencyという。ただし空間周波数は,縞模様だけに適用されるのではなく,視野内の特定の方位軸に沿った変調が何周期あるかを記述する一般的な概念である。
縞視力の測定を拡張し,正弦波の輝度変調が検出されるために必要な最小の輝度コントラストを空間周波数の関数として測定したならば,得られた曲線の高域カットオフ周波数が縞視力に相当し,曲線全体は空間的コントラスト感度関数spatial contrast sensitivity functionとよばれる。平均輝度・刺激サイズ・網膜部位などにより異なるが,明所視ではこの関数は中心視でおおむね3~5cpd(cycles per degree)をピークとするバンドパス型の逆U字型曲線となり,視野周辺に行くにつれてピーク周波数が低域にシフトする。暗所視では低周波側で感度の落ちないローパス型となる。空間的コントラスト感度関数をもたらすメカニズムは単一でなく,限られた通過帯域をもち,最適周波数の異なる複数の空間周波数チャンネルspatial-frequency channelが介在すると考えられている。
信号雑音比が悪く,欠損値も生じる貧しい視覚入力から可能な限り有意味な情報表現を構築するために,視覚系では特徴的な空間処理がさまざま行なわれており,それらを反映したさまざまな視覚現象が報告されている。その一つが明るさ・色の同時対比simultaneous contrastであり,同じ輝度の面であっても暗い面に囲まれた場合はより明るく,明るい面に囲まれた場合はより暗く感じられる。肌理・運動・奥行きの属性においても対比は生じる。この対比作用には,空間変化を強調する働きが関係すると考えられている。もう一つの例が知覚的充塡perceptual filling-inであり,輪郭がはっきりしない領域において,領域の周囲にある明るさや色などが領域内部全体に侵蝕するように感じる。盲点での充塡はその典型例だが,それ以外にもトロクスラー効果Troxler effectという名称でよばれるようにぼやけた輪郭を視野周辺で眺めつづけた場合などにも生じ,視覚系の一般的特性といえる。肌理・運動・奥行きの充塡も報告されている。
刺激画像の物理的構造とそれを観察したときの見かけが一致しない現象を指して,錯視visual illusionという。古来より数多くの幾何学的錯視geometrical illusionが報告されており,網膜像とわたしたちの長さ・大きさなどの種々の知覚が一致しない例は枚挙にいとまがない。錯視図形を観察している際も,われわれの視覚情報処理系では,貧しい入力から可能な限り外界構造を正しく推定しようという計算回路が働いているはずである。そして,幾何学的錯視をはじめとする数多くの錯視現象は,入力と出力との関係が特異であることから,そのような計算回路の原理にヒントを与えてくれる有効な実験材料である。
【時間特性】 時間的に輝度を変調させて,時間的コントラスト感度関数temporal contrast sensitivity functionを測定することができる。平均輝度・刺激サイズ・網膜部位などにより異なるが,この関数の形状はおおむね5~15㎐をピークとするバンドパス型の逆U字型曲線を呈するが,高域カットオフ周波数はおおむね50~60㎐であって,これを臨界ちらつき頻度critical flicker frequencyという。輝度を変えず色情報のみを変調させた場合はローパス型に変わり,色の臨界ちらつき頻度color critical flicker frequencyは約15㎐となる。
空間的な同時対比があるように,時間的にも明るさや色の継時対比successive contrastという一過性の現象があり,同じ灰色でも,同一領域に暗い刺激が呈示されていた後では明るく見え,明るい刺激が呈示されていた後では暗く見える。このように以前の刺激と逆の側に知覚がバイアスされることを一般に陰性残効negative aftereffectといい,明るさや色だけでなく方位の傾きや運動など多種の属性において生じることが知られ,特異的な残効が生じることをもって,その属性を特異的に処理するメカニズムが存在する証拠とされる。
視覚情報から時間そのものを認識することに関しては,二つの刺激を呈示して同時性判断や時間順序判断を行なう研究例があるが,2刺激の時間差が両眼視差や運動のメカニズムによって検出されるような刺激呈示事態でない限り,聴覚に比べて視覚の時間差の弁別能ははるかに低い。時間間隔の知覚に関しては聴覚刺激と同様,変化が豊かに含まれる刺激は知覚的に長く感じるなど,多感覚にわたり共通した特徴がある。
【注意と意識】 網膜に像が投影され,初期の神経メカニズムによって処理がなされたとしても,つねに意識体験として視覚像が感じられるとは限らない。両眼視野闘争binocular rivalryはその好例だが,その極端な形としては,一方の眼に動的に切り替わる刺激を呈示しつづけると,他方の眼に呈示された刺激が意識に上らない状態が長時間持続し,これを連続フラッシュ抑制continuous flash suppressionという。また,両眼分離視に限らず一般に,注意を向けやすい刺激が存在する状況下ではそれ以外の対象の存在や,対象の変化が意識化されにくくなる。これらの現象はそれぞれ非注意による見落としinattentional blindness,チェンジ・ブラインドネスchange blindnessとよばれている。多義図形における図と地の反転と同じく,同じ網膜像が入力されていながら視覚対象が意識化されるか否かが変わるという現象は,脳内の状態の切り替わりを反映しているはずであり,このような事態は意識の神経相関を実証的に探究するうえで有用な研究材料である。
【多感覚相互作用】 視覚と身体運動感覚には密接なかかわりがある。たとえば,頭部が日常的につねに揺れ動いているにもかかわらず,頭部回転を補償する作用のある前庭動眼反射vestibulo-ocular reflexという眼球運動によって,ある程度安定的な網膜像が得られている。頭部に対して固定的な位置に置かれた眼前の固視点を見つめながら頭部回転をすると,反射は抑制されるが,この場合は固視点が回転方向側に動いているような眼球旋回錯視oculogyral illusionが生じる。視覚情報から身体運動感覚が惹起される例としては,視野の広い領域に動きがあると反対方向に自分の身体が動いていくようなベクションvectionという錯覚を感じる。また,ベクションが生じるのに伴い立位において重心動揺が生じることがある。ベクションに対抗して身体重心を移動すると姿勢安定が当然損なわれるため,前庭器官や自己受容感覚器からの信号で重心の再補正が行なわれ,この反復が重心動揺となると考えられる。
音声の音像定位が話者の映像の位置にバイアスされて錯誤される腹話術効果ventriloquism effectは,聴覚が視覚に影響されるビジュアル・キャプチャーvisual captureと総称される多感覚相互作用の好例だが,視覚体験もまた他の感覚モダリティの刺激の存在によって左右される。たとえば二重フラッシュ錯覚double-flash illusionという現象では,フラッシュ光を1回だけ点灯し,それとほぼ同時にクリック音を2回続けて呈示すると,光が2回点灯したように錯覚される。これは時間解像度に優れた聴覚信号に依存して,視覚対象の時間知覚が変容する例である。また,運動方向が曖昧な図形を観察する場合,自分の腕をある方向に動かすと,図形の運動方向が腕の運動方向に引っ張られて見えることがある。
【神経メカニズム】 このような多様な視覚機能は,精緻な生体メカニズムに支えられている。外界からの光学像は眼の角膜corneaで粗な屈折を受け,水晶体lensにおいて屈折の微調整を受けた後,透明なゲル状組織である硝子体vitreous bodyを通過して網膜に結像する。水晶体は毛様体筋の力で屈折率を適応的に変化させる。瞳孔径は瞳孔括約筋・瞳孔散大筋により径が適応的に変化し,対光反応をはじめとする種々の原因で生じる縮瞳は,入射光量を減ずるとともに焦点深度を増す機能をもつ。網膜の最外層である外顆粒層・桿体錐体層にある視細胞photoreceptorにおいて,受光量の増加に対して細胞膜が過分極応答する形で光電変換される。さらにその外の層にはメラニン色素に富む色素上皮が接し,乱反射を防いでいる。
視細胞外節には多数の膜が重なっていて,ビタミンAの一種のレチナールを含んだタンパク質である光感受性分子がそれぞれの膜に含まれている。この分子を視物質photopigmentとよぶ。光量増加に対して視物質が構造変化して連鎖的な生化学反応を引き起こした結果,細胞膜ナトリウムチャンネルの閉確率を高めて膜電位を過分極側に変化させる。視細胞には大きく分けて明所で働く錐体coneと暗所で働く桿体rodの2種類がある。錐体にはL錐体,M錐体,S錐体があり,各々の視物質は光の波長に対する感度(分光感度)が異なっていて,それぞれ560㎚,530㎚,425㎚に最大感度をもち,これら3種類の錐体が明所視photopic visionにおける色覚の最初の神経基盤となっている。桿体の視物質は495㎚に最大感度をもつ1種類しかないため,暗所視scotopic visionにおいては色は見えないが,光電変換効率はきわめて高い。すなわち,1個の視物質においてレチナールが11-シス型の状態から光エネルギー吸収後に立体異性化して全トランス型に変わり,これに伴いタンパク質がロドプシンからメタロドプシンへと構造変化することで,細胞内液にある数百個のGタンパク(トランスデューシン)を活性化し,その各々が1個のホスホジエステラーゼを活性化,その各々が膜チャンネルに作用するサイクリックGMPを数百分子/秒の速度で分解するという反応連鎖を経て信号が増幅する(カスケード増幅)。このため,1個の光子が視物質に当たることが膜電位変化を引き起こしうる。明所視と暗所視との境目を薄明視mesopic visionといい,錐体と桿体の両方が寄与する。
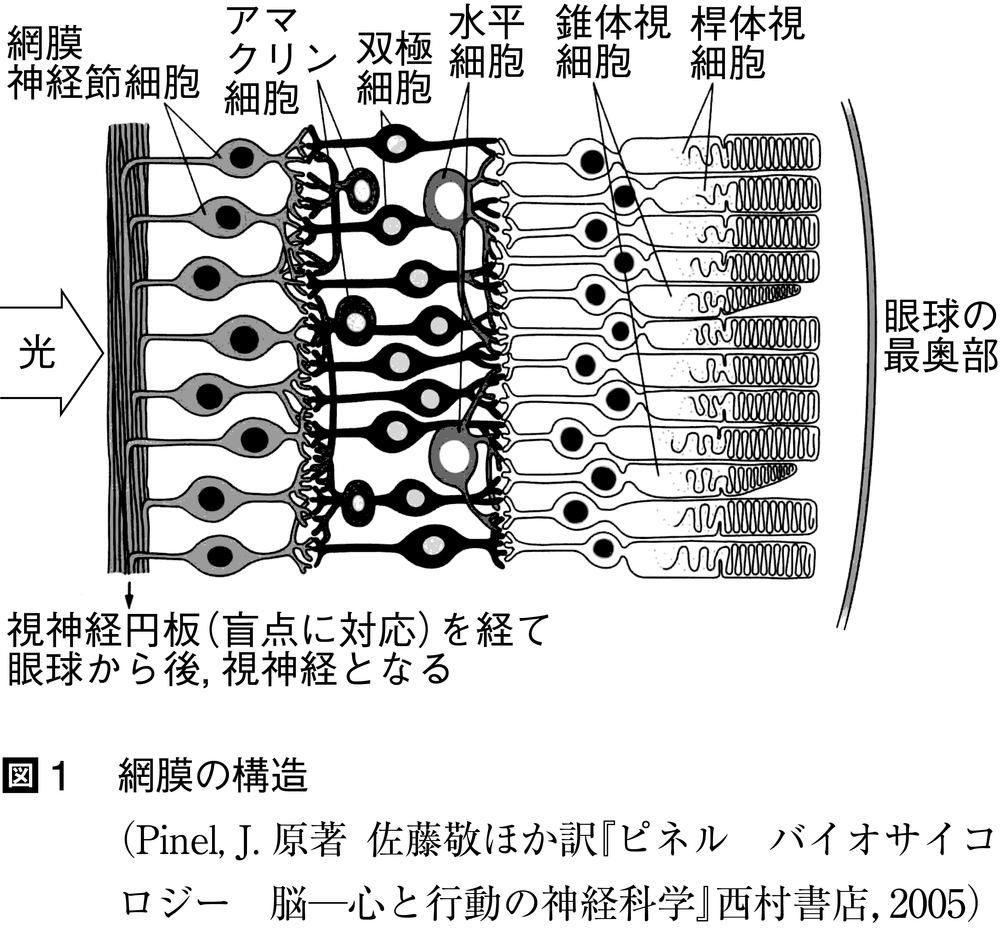
視細胞の膜電位変化はシナプスにおける神経伝達物質の放出量の変化をもたらし,次の双極細胞bipolar cellの膜電位変化をもたらす。視細胞と異なり,双極細胞においては,光量増加に過分極応答するOFF型と脱分極応答するON型が出現する。これらの膜電位変化はシナプスを介して次の網膜神経節細胞retinal ganglion cellに興奮をもたらし,活動電位の発生確率を変化させるが,ここでも暗い光に対して興奮性に応答するOFF型と明るい光に対して興奮性に応答するON型がある。これらの直列的伝達経路は介在ニューロンによる修飾も受け,隣り合う視細胞や双極細胞は水平細胞horizontal cellによって,隣り合う双極細胞や網膜神経節細胞はアマクリン細胞amacrine cellによって,それぞれ水平方向の結合をしている(図1)。
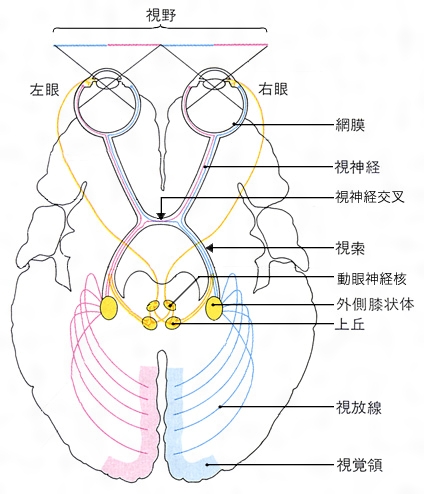
網膜神経節細胞の軸索は視神経円板を貫通し視神経束となって眼から脳へと伸び,視交叉で半交叉(網膜耳側・鼻側のニューロンがそれぞれ同側・対側の大脳半球へ連絡すること)した後は視索とよび名が変わり,間脳にある視床の外側膝状体lateral geniculate nucleus(LGN)に終わるものと,中脳の上丘を経て視床枕pulvinarに連絡するものがある。前者が視放線を介して大脳皮質の一次視覚野primary visual area(V1野)に連絡し,意識的視覚にかかわる脳内情報処理に寄与するのに対し,後者は皮質下核と連絡をもち,眼球運動や種々の視覚性反射を主に取りもつとされている。
霊長類の網膜神経節細胞は,時間解像度が高く空間解像度が低いパラソル細胞と,空間解像度が高く時間解像度が低いミジェット細胞と,その他の細胞に分類される。パラソル細胞はLGNの大細胞層,ミジェット細胞は小細胞層に投射し,またこれらの層の間にある顆粒細胞層へ投射する細胞がある。ごく一部の網膜神経節細胞には光感受性があり,光を直接受容して活動電位を発生する。この内因性光感受性網膜神経節細胞intrinsically photosensitive retinal ganglion cell(ipRGC)の送る信号は視交叉上核へ送られ,概日リズムを形成する視覚性同調因子と考えられたが,ほかの視覚機能にも関与している可能性が示唆されている。
LGNの大細胞層からはV1野の4Cα層,小細胞層からは4Cβ層,顆粒細胞層からは1~3層に投射がある。V1野以降は,視覚前野と総称される多くの皮質領野(V2,V3,V4,etc.)で階層的に情報処理が進んでいく。大細胞系の神経連絡は視覚前野のMT野,MST野,LIP野などから成る背側経路dorsal pathway(ミシュキンMishkin,M.の分類でいう「where」経路)に連なり,運動・奥行き・空間定位に関与する。これに対し,小細胞系の神経連絡はV4野,下側頭皮質(IT),ヒト脳ではさらにLOC,FFA,PPA,EBAなどの区分のなされる腹側経路ventral pathway(「what」経路)に連なり,色・立体形状・物体認識に関与する(248ページ図2)。これら2経路の間には相互連絡があり,また各経路内でも上位と下位の領野同士には双方向結合がある。各々の半視野は対側の大脳半球で表現されるが,両半球を連絡する脳梁を介して互いの半視野表現の間に情報のやりとりがある。色や運動など異なる感覚属性の処理が異なる神経回路でなされるなら,それらの処理結果をどうにかして結びつけて合一な対象認識に至る必要がある。神経線維で相互連絡された神経ネットワークが同期的に振動する状態が,この結合問題binding problemの解決の役目を担っているという考え方がある。
視覚ニューロンの各々は受容野receptive fieldをもち,視野内の一部の領域の光刺激に対してしか応答しない。多くのミジェット細胞の受容野は同心円型で,受容野中心部は光オンで興奮し,周辺部は光オフで興奮する(中心-周辺拮抗型)。あるいはこれと逆の極性の受容野をもつといったように,受容野中心部と周辺部で応答特性が異なる。持続する光強度に対する持続的な応答成分とは別に,光量の時間的変化に対する一過性の応答成分がある。大脳皮質の視覚ニューロンは,より複雑な受容野構造をもつ。方位選択性ニューロンは,特定の傾きをもつ光刺激,方向選択性ニューロンは特定の向きに動く光刺激,色選択性ニューロンは特定の色,視差選択性ニューロンは特定の両眼視差に対して選好応答する。高次領野になるにつれて,刺激に対する選択性は複雑性を増す。たとえば,手のような形にしか応答しないサルIT(下側頭領域)のニューロンが見つかっているし,サルITやヒトFFA(紡錘状回顔領域)では顔刺激に対して選択的に活動が増す。
【視覚異常】 網膜上に正しく結像することを正視といい,それより近位・遠位に焦点がある屈折異常をそれぞれ近視・遠視という。老眼とは,加齢により水晶体の弾力が落ちて近見時に像がぼけることを指す。これらの屈折異常が光学矯正により視力を回復できるのに対し,弱視amblyopiaは斜視,屈折の左右差,発達時の視覚遮断などの原因で生じ,異常眼で視力が低い状態で,眼光学系に原因が求められない。この弱視を含め,視機能が低く社会生活を営むのに不自由があり,光学矯正で容易に回復できない障害をロービジョンlow visionと総称する。視野狭窄や暗点など,網膜や眼内の黄斑色素の加齢による変性との関連が疑われる症状で悩む事例も多く,とくに現代の高齢化社会で問題視されている。
視覚神経系の機能不全により起こる視覚性認知の障害も数多く報告されている。色の認知だけが損なわれる大脳性色覚障害や,物の動きだけがわからない障害がある。形態認知の障害では,何の絵かわからない画像失認,どんな実物体かわからない物体失認,だれの顔かわからない相貌失認,どの街かわからない街並失認,読解や音読ができない失読がある。空間認知の障害では,絵が描けない構成障害,目標に向けて手を到達運動できない視覚運動性失調,見慣れた場所で道に迷う地誌的失見当などが挙げられる。注意の障害では,左右半視野に二つの物体を同時呈示すると一方(多くは左側)を無視してしまう消去,視野や対象の片側(多くは左側)を無視してしまう半側空間無視,複数対象に同時に注意を払えない背側性同時失認,複数対象の全体的意味を瞬時にとらえられない腹側性同時失認などがある。また,失認とは異なり対象を誤って認知してしまう症状には,対象が消失しても視野内にありつづけるように見える視覚性保続,存在しない光や物が見える幻視,物の形が歪んで見える変形視などがある。これらの神経心理学的症状と大脳皮質領野との関連性が調べられている。 →明るさの知覚 →色 →感覚 →眼球運動 →恒常現象 →錯覚 →視覚刺激 →視覚領野 →視野 →順応 →知覚 →両眼視
〔村上 郁也〕

出典 最新 心理学事典最新 心理学事典について 情報
改訂新版 世界大百科事典 「視覚」の意味・わかりやすい解説
視覚 (しかく)
visual sensation
光刺激によって生じる感覚で,明暗を感じる感覚を光覚,色を感じる感覚を色覚という。おもな感覚器は目である。原始的な光覚は神経光覚器や皮膚光覚器によって起こる。よく発達した視覚は聴覚や嗅覚と同様に,動物が遠くはなれた所からの情報を得るのに重要な感覚である。ことに外部の情況を立体的に知るうえでは視覚が最も優れている。視覚機能は明暗識別や光源の方向識別,あるいは対象物の形態や動き,対象物までの距離などの識別や色彩弁別に分けられる。
原始的な神経光覚や皮膚光覚では明暗の識別ができる。例えば,オオノガイのような二枚貝は,水管に影がさすと水管を反射的に引っ込める。影にのみ応答するので陰影反射と呼ばれている。原始的光覚器は脊椎動物にもある。例えば,魚類や両生類,爬虫類の脳の松果体は,光の明暗に応答する光覚器である(顱頂眼(ろちようがん))。光がくる方向に,またはその逆方向に定位して動物が動くことを光走性というが,光走性は原始的な光覚器をもつ動物から,発達した目をもつ動物までみられる一般的な視覚行動である。例えば,ミミズでは光走性は,皮膚光覚器により明暗を手がかりに行われるが,昆虫では,複眼により光源を網膜にとらえたり,左右の目に入る光の明るさが等しくなるように動いて光源に定位する。
発達した視覚器としては節足動物の複眼や脊椎動物と軟体動物頭足類のカメラ眼があげられる。これらの目は比較的よい解像力をもち,形の識別,動きの識別などに適している。例外はあるが,解像力は一般に体高の高い動物の方がよい。視力1.0のヒトの目の分解能は0.01度であるが,ネコでは約0.1度,トンボでは約1度,ハエでは約2度である。光点の点滅を識別できる限度を点滅の頻度で表したものをフリッカー融合頻度(ちらつき融合頻度)というが,この頻度が高い方が動きの識別能力が高い。ヒトのフリッカー融合頻度は70~100Hzであるが,ハトでは150Hzと高く,また速く飛ぶ昆虫ではフリッカー融合頻度はひじょうに高く,例えばハエでは約300Hzである。したがって,昆虫は図形識別能力は悪いが,動きの識別は格段によいことになる。また,脊椎動物の霊長類や鳥類,魚類などや,昆虫類には色覚が発達した動物がある。
動物の視細胞visual cellは光で刺激される部分が特別な構造をしている。脊椎動物の視細胞の光刺激を受容する部分は外節と呼ばれ,膜が重なり合った構造になっていて,この膜に感光色素が配列している。外節は繊毛の変形したものである。脊椎動物の視細胞は,外節の形の違いにより錐体と杆体(かんたい)に分けられる。錐体は明るいときの視覚(昼間視)に関係し,杆体は薄暗いときの視覚(薄明視)に関係する。錐体は刺激となる光の波長の違いで3種類のものが区別され,色覚に関係する。
無脊椎動物の視細胞において光刺激を受容する部分は,微絨毛が突き出して密生した構造のものが多く,繊毛が変形したものは例外的にしか知られていない。複眼では個眼に数個の視細胞があり,各視細胞は個眼の軸の周りに微絨毛を突き出すので,個眼の中心には軸に沿って細長い杆状の微絨毛の集合物ができる。これを感杆と呼び,このような視細胞を感杆型の視細胞と呼び,脊椎動物の繊毛型の視細胞と区別することがある。
視細胞に光が当たったときに,光のエネルギーを吸収して光化学変化を起こすのが感光色素である。杆体の感光色素はロドプシンで,光が当たると分解してビタミンA1のアルデヒドであるレチナールとオプシンというタンパク質になる。暗黒中では,十分レチナールが存在するときには,レチナールとオプシンからロドプシンが再生する。錐体の感光色素はアイオドプシンである。昆虫や軟体動物頭足類の感光色素もロドプシンであるが,脊椎動物のロドプシンとは異なり,完全に分解せずにメタロドプシンに変わるだけである。
光刺激により視細胞には膜電位変化が起こる。脊椎動物の網膜には双極細胞や水平細胞,アマクリン細胞などの介在神経があり,視細胞に起こった変化はこれらに伝えられ,最後に視神経のインパルスとなり,大脳皮質に伝わる。光刺激に対する視神経繊維の応答の特徴の一つは,側抑制がみられることである。側抑制は,最初カブトガニの複眼の視神経繊維で観察されたもので,一つの個眼を光刺激すると,刺激された個眼の視神経繊維にインパルスが発生すると同時に,隣接する個眼の視神経繊維には興奮が起こりにくいように抑制がかかる現象をいう。側抑制は明暗の境界線での対比を強調するなどの働きをする。
執筆者:立田 栄光
視覚の生理学
ヒトの視覚に目が必要なことは万人の知るところであるが,目は視覚の入口にすぎない。物の映像は目によってとらえられ,視覚信号に変換され,視神経によって脳に送られ,これが脳によって処理されて初めて視覚が成立するのである。このような視覚認知の過程はコンピューターによる文字読取りの働きと対比される。文字読取りの場合は,文字の映像がテレビカメラによってとらえられ,これが電気信号に変換され,コンピューターにより分析されて,文字が判読される。文字の読取りは,テレビカメラによる光学的な図形情報から,電気的な図形情報への変換と,コンピューターによる電気的な図形情報の解読の二つの過程をふんで成立する。視覚認知もまったく同じようにこの二つの過程からなる。第1の光学的な視覚情報から,電気的な視覚信号への変換が目の網膜で行われ,第2の視覚信号の解読が大脳で行われるのである。
視覚の情報処理
外界の映像は目のレンズ(水晶体)を通して,網膜に投影される。網膜には数億個の神経細胞があり,ここで光学的な映像が電気信号に変換される。この電気信号が100万本の視神経繊維によって大脳の視覚中枢に送られる。1本1本の視神経はいわばテレビの画像の画素にあたり,その反応の強さは画素の明るさを表す。視神経の数が多ければそれだけ精細な映像信号を大脳に送ることができる。網膜は単に光学的な映像を忠実に電気信号に写像するだけでなく,映像の輪郭などの視覚認知に欠くことのできない重要な成分をとくに強調して大脳に送る。この働きは側抑制と呼ばれる神経機構によって行われる。
大脳の視覚中枢(視覚野ともいう)は左右の大脳半球の後部(後頭葉)にある。両眼から出た視神経繊維は,視交叉部で内側の視野からくる半数が交叉(これを半交叉という)し,交叉しないそれぞれの外側の繊維とともに,左右の後頭葉へと導かれる。この結果,右側の視野の視覚情報は左側の大脳で,左側の視野の情報は右側の大脳で処理される(これを〈対側支配の原則〉という)。
大脳の視覚中枢には約10億の神経細胞がある。これらの細胞はそれぞれ特有の図形にだけ反応する。この性質は反応選択性と呼ばれ,大脳の視覚細胞の働きを左右する最も重要な特性である。図形が示されたときにある細胞が反応すれば,その細胞が好んで反応する特徴がその図形に含まれていることを意味する。だから細胞は図形の特徴をとり出す特徴フィルターの働きをもつ(これを特徴抽出機能という)といえる。視覚中枢では図形は特徴の集りとして認知される。網膜では図形が,さまざまの明るさをもつ点の集りとして認知されるのと対照的である。
視覚中枢は特徴抽出機能に基づいて第一次視覚野と高次視覚野に区分される。網膜から送られてくる視覚情報はまず第一次視覚野で分析され,その結果が高次視覚野に送られ,さらに,高次の視覚情報がとり出される。
第一次視覚野
第一次視覚野には直線の位置,幅,傾き,角,運動方向,黒白あるいは色コントラストなどの特徴(これらを図形の幾何学的パラメーターという)に反応する細胞がある。神経細胞はこれらのいくつかのパラメーターに対し,反応選択性を備え,その条件がすべて満足されたときに初めて反応する。これらの細胞により,すべての図形は直線成分に分解され,その幾何学的特徴(例えば正方形の辺,角,傾き,位置)や図形の運動方向,左右の目でとらえられる位置のずれ(視差),色(光の波長)などの視覚情報がとり出される。
高次視覚野
第一次視覚野が視覚神経から送られてくるあらゆる視覚情報を扱うのに対し,高次視覚野はいくつかに区分され,ある視覚情報を専業的に分析する。例えば,色の認知を担当する高次視覚野(ここを色中枢という)では,すべての細胞が色に対して反応選択性をもつが,他の特徴に対してはほとんど反応選択性がない。ここの視覚細胞はどのような光の下でも,色を正しく判断する働き(これを色恒常性という)をもつ。青みがかった光で赤い色紙を照らせば,赤い色紙から反射される光は青い光の成分を強く含む。逆に赤みがかった光の下では赤い成分の光が強く反射される。このような条件の下でも色中枢の細胞の色紙に対する反応性は変わらない。赤色に反応する細胞はどちらの条件でも赤い色紙に反応する。第一次視覚野の赤細胞が赤い光で照らされた青い色紙に反応し,青い光で照らされた赤い色紙に反応しないのと対照的である。第一次の視覚野は色紙から反射されてくる光のスペクトル成分に対してのみ反応選択性をもつ。だから赤細胞は赤い光のスペクトル成分が強ければ,色紙の色とは関係なく反応するのである。しかし色中枢の細胞は,見ているものから反射される光の赤成分の相対的スペクトル強度に対して,反応選択性をもつ。そこで赤細胞は,見ているものから反射された光の赤成分のスペクトル強度が,周りのものから反射されたものよりも相対的に強ければ,赤成分のスペクトル強度とは関係なく反応するのである。この反応選択性は,いくつかの第一次視覚野の細胞で分析された光のスペクトル成分の情報を総合して得られると考えられる。
三次元の動きも高次視覚中枢によって専業的に分析される。前後方向の動きは次のような二つの視覚的手がかりを使って認知される。一つは,物の動きを両眼の網膜上の動きの違いから認知する方法である。すなわち,物が鼻に向かって近づくときには右目の網膜の映像は右方向,左目では左方向に動き,遠ざかるときには左目では右方向,右目では左方向の動きが生じる(これを運動視差という)。等距離で運動するときにはこのような運動視差は生じない。いま一つは,像の大きさから検知するもので,物が近づくときには網膜の映像の大きさは距離に反比例して大きくなる。運動認知の中枢にはこの二つの視覚的手がかりの組合せに対して反応選択性を持っていて,前後方向の動きを認知する細胞がある。例えば接近運動を認知する細胞は右目に右方向,左目に左方向の運動視差と映像が大きくなるという刺激が組み合わさって生じたときに反応する。言い換えれば,この二つの視覚的手がかりが生じたときには,そのものは近づいてくると判断するわけである。左右の目でとらえられた映像の動きや輪郭の拡大や縮小が第一次の視覚野で抽出され,運動認知の中枢はそれらの情報を総合して三次元の動きという特徴をとり出すのである。
視覚中枢の可塑性
このような特徴抽出機能は単一の視覚細胞の属性として備わっているのでなく,いくつかの神経細胞がシナプスを介して互いに結合し神経回路をつくることから生まれる。コンピューターの計算素子であるトランジスターそれ自身が計算能力をもつのでなく,いくつかのトランジスターが回路をつくることによって計算能力が生じるのと同じである。そこで視覚野には互いに似かよった特徴抽出機能をもつ細胞が柱状に連なって配列されている。互いに似た働きをもつ細胞が近くにあれば,回路を形成するのに便利だからである。
これらの神経回路は生得的なものではない。生後のある時期(ヒトでは2~6歳まで)に形成される。生まれたての動物の視覚細胞を調べると,成長した動物で見られるような反応選択性がまったくない。この時期の視覚細胞はすべてのものに無差別に反応する。また,この時期に両目を縫い合わせて視覚体験を奪われた動物は,その後,目をあけても視覚は永久に回復することなく,視覚細胞の反応選択性はほとんど失われている。このことから生後しばらくの間は,神経細胞をつなぐシナプスが変わりやすく(可塑的),そのときに見たものに対して最もよく反応するように,シナプスのつながりが変わる,つまり視覚体験を学習することによって,反応選択性が後天的に獲得されると考えられる。
われわれの脳にはそれぞれの視覚認知に特有の神経細胞があり,それが活動したときにそれに対応するものが外の世界に存在すると判断する。神経細胞の活動という脳の内なる世界のできごとを通して,外の視覚世界をうかがい知るのである。この内なる世界は生まれつき存在するのではなく,生後のある期間の視覚体験によって形成される。すなわち,後天的に獲得された内なる世界を通して外の世界が認知されるのである。
→感覚 →目
執筆者:外山 敬介
出典 株式会社平凡社「改訂新版 世界大百科事典」改訂新版 世界大百科事典について 情報
日本大百科全書(ニッポニカ) 「視覚」の意味・わかりやすい解説
視覚
しかく
可視光線の刺激によっておこる感覚をいい、これによって外界の事物や現象が認知される。視覚は聴覚、味覚、嗅覚(きゅうかく)などとともに特殊感覚の一つとされる。
[市岡正道]
目の通光学
視覚の受容器細胞は眼球の内面を覆っている網膜にあり、網膜中の視細胞(錐状体(すいじょうたい)と桿状体(かんじょうたい))によって光の刺激を感受する。このため、他の眼球を構成する組織は、物体の像を正しく網膜に結ばせるための通光学的装置といえる。光線を屈折させる装置を外側からあげると、涙液、角膜、前眼房水、水晶体、および硝子体(しょうしたい)の五つがある。このうち水晶体の屈折率がもっとも大きく、光の屈折はおもに水晶体を通して行われている。正常眼では、眼前約6メートル以上の距離にある物体は屈折させることなく網膜上に正しく結像するが、これより手前にある物体は網膜の後方に像を結ぶため、屈折力を増加させなくては正しく結像しない。このように、近い物を明視する(はっきりと見る)ために目の屈折力を増すことを「目の遠近順応」という。無順応眼(遠近順応を行わない目)で明視できるもっとも遠くにある点を遠点、最大に遠近順応を行った目で明視できるもっとも近くにある点を近点という。近点以内の物体は明視できない。
屈折力の増大は水晶体を厚くすること(とくにその前面の彎曲(わんきょく)度を増すこと)によってもたらされるが、年齢とともに水晶体が硬化して肥厚しにくくなると、遠近順応力もそれに伴って減退してくる。したがって、近点は年齢とともに遠ざかることになる。たとえば、20歳の近点は眼前約10センチメートルであるが、40歳では約20センチメートル、60歳では約80センチメートルに遠ざかる。このように40歳代以降になって近点が遠ざかることを老視(老眼)という。また、眼球の前後軸が正常より長い目では、平行光線が網膜より前方で結像する。これを近視(近眼)といい、凹レンズで矯正する。これに対して、眼球の前後軸が正常より短すぎる目では網膜より後方で結像する。これを遠視といい、凸レンズで矯正する。また角膜の曲率が一様でない場合には屈折度も異なるため、角膜のある経線上の光線と別の経線上の光線は別々の深さで結像し、像はぼやけて見える。これを乱視という。
[市岡正道]
網膜の働き
物体の像を結ぶ網膜は10層の細胞層からなり、視細胞である桿状体(桿体)と錐状体(錐体)とは最外層に位置している。桿状体は網膜の周辺部に多く分布し、光に対する感受性が高く、暗い所で明暗の差を感知できる。錐状体は網膜の中心部に多く分布し、光に対して閾値(いきち)(感覚をおこすに有効な最小値)が高く、明るい所で色を感知する。両者にはそれぞれ特有の感光性色素が含まれており、光が当たると化学変化をおこす。これが「視細胞の興奮」である。
この両視細胞は、双極細胞を介して神経細胞に連なっている。神経細胞の軸索(神経細胞より出る長い突起)は眼球を出ると視神経となり、視神経交叉(こうさ)を経て視索に続き、外側膝状体(しつじょうたい)に達する。視神経交叉では、両眼の網膜の左半分に由来する視神経線維は左側の、右半分に由来する視神経線維は右側の膝状体に達している。外側膝状体から発する視神経線維は視放線となり、同側の後頭葉の視覚領に終わる。視神経の軸索の枝は視索から離れて上丘に達し、視覚に基づくいろいろな反射(たとえば瞳孔(どうこう)反射や眼球運動反射など)を行っている。
なお網膜は、細動脈が直接観察できる身体唯一の場所である。このため、糖尿病、高血圧、その他の血管の疾患の診断や病状判定には、検眼鏡による網膜の細動脈観察が重要な意味をもっている。
[市岡正道]
明暗感覚と色覚
視覚には、光の量の多少によって生ずる明暗感覚と、光の波長の差に基づく色覚の二つがある。明暗感覚は外界の明るさによって変化するものであり、たとえば明るい所から暗い所へ入ると初めは物が見えないが、しだいに網膜の光に対する感受性が高まり、10~20分もすると物が見えるようになる。これを暗順応という。これに対して、暗い所から明るい所に出ると初めはまぶしくて物が見えないが、やがて網膜の光に対する閾値が高まってきて、目はその明るさに慣れてくる。これを明順応といい、明順応には約3分間を要する。暗順応は最初は網膜中心部にある錐状体を介して行われるが、この順応は軽度であり、これに続く網膜周辺部の桿状体における暗順応のほうが強く、長続きする。
明るさの感覚(明度)は、光の波長、すなわち色(色相)によっても異なる。明るい所では黄(550ナノメートル付近)がもっとも明るく見え、スペクトルの長短両端へ向かうにつれて暗く見える。しかし光度を弱めていくと、もっとも明るく見える部位が黄から青緑(510ナノメートル付近)に移る。これを「プルキンエ現象」Purkinje phenomenonといい、1825年に発見された。夕暮れや明け方といった薄暗いときに緑や青が明るく際だって見えるのは、この現象のよい例である。
[市岡正道]
視知覚
まっすぐ前方を向いていて目に見える範囲を視野といい、眼球を動かして見える範囲を注視野という。視力は、それぞれある大きさ、あるいはある広がりをもった2光体を2光体として分離識別できる最小視角の逆数をもって表される。実際には、6メートルの距離からの最小視角が1分(ぷん)のときを視力1としている。このときの2光体の網膜内の像の隔たりは約4マイクロメートルで、これは錐状体1個の幅にほぼ匹敵する。視力は複雑な要因で決められるものであって、光の照射方法、光の明るさ、光と背景との対比などのほか、眼球での結像の仕方、錐状体の性状などによっても影響される。
繰り返し与えられる点滅光刺激が一つ一つの光刺激として感じられる最大点滅頻度を臨界融合頻度、またはフリッカー値という。中等度の明るさの光刺激のときのフリッカー値は毎秒35~70である。フリッカー値は、二つの色光の明るさの比較や疲労度の測定などに応用されている。
物体の奥行を知覚することは、単眼でも両眼でも可能である。単眼視の場合は、遠近に対する遠近順応の努力の差や視差(目または物体が移動することにより網膜上の像が移動すること)によって奥行が知覚される。また経験によって物体の大きさや形などを知っている場合には、陰影のでき方などによって奥行が知覚される。両眼視の場合は、主として輻輳(ふくそう)度(両眼注視線が見ようとする一点に集合する度合い)と両眼視差(両眼網膜像の差)によって知覚されるが、単眼視の場合に比べれば一段と精密である。
[市岡正道]
動物の視覚
ヒトに限らず、動物一般についても、光によって生ずる感覚を視覚という。通常は、広義の光感覚と区別し、特定の光受容器である目の働きによる感覚をさすが、光感覚と同義に用いられることも多い。視細胞に光が当たると、細胞に含まれる感光色素タンパク質(視物質)の光化学作用によって視細胞に電気的な応答が引き起こされる。脊椎(せきつい)動物の視細胞では、網膜の桿状体細胞と錐状体細胞に生ずる変化は、膜を横切って流れるナトリウム電流の低下による過分極であるが、無脊椎動物における変化は脱分極である。視覚は主として目の構造に応じて発達し、明暗、光のくる方向、物体の動き、形や遠近などが識別されるようになる。また異なる波長の光を吸収する視物質の組合せによって波長の差が識別される。脊椎動物や棘皮(きょくひ)動物の視細胞は、繊毛軸糸をもった繊毛型であるが、節足動物や軟体動物の視細胞は、微絨毛(びじゅうもう)の配列する感桿型である。節足動物では多くの個眼が集合した複眼をもつが、軟体動物と脊椎動物は発達したカメラ眼による形態視を行う。
[村上 彰]
[参照項目] | | | | | | | | |
百科事典マイペディア 「視覚」の意味・わかりやすい解説
視覚【しかく】
→関連項目盲人
出典 株式会社平凡社百科事典マイペディアについて 情報
ブリタニカ国際大百科事典 小項目事典 「視覚」の意味・わかりやすい解説
視覚
しかく
vision
出典 ブリタニカ国際大百科事典 小項目事典ブリタニカ国際大百科事典 小項目事典について 情報
普及版 字通 「視覚」の読み・字形・画数・意味
出典 平凡社「普及版 字通」普及版 字通について 情報
栄養・生化学辞典 「視覚」の解説
視覚
世界大百科事典(旧版)内の視覚の言及
【光学】より
…このような意味での今日の光学は物理学の一分科にすぎないが,少なくとも17世紀までは,科学者ばかりか,哲学者や神学者さえもこの学問に取り組んだ。しかも,光学の名のもとに,反射や屈折だけではなく,視覚の問題や,場合によっては眼球の解剖学と生理学すら論じられたのである。 まず,古代には,数学者として知られるユークリッド(エウクレイデス)が光の直進性と反射を研究し,哲学者のアリストテレスも色彩の問題を論じた。…
【心身問題】より
…とりわけ外部刺激が脳に作用して世界風景が見え聞こえるという生理学公認の事実の再解釈が必要である。視覚を例にとる。まず視覚の風景が〈見透し〉構造をもつことに留意する。…
【知覚】より
…ケーラーは,あらゆる知覚現象には必ずそれに対応する脳の生理的過程があるという心理物理同型論psychophysical isomorphismの立場から,仮現運動が実際の運動と等しい生理過程を大脳皮質にひき起こすのであろうと考えた。最近の神経生理学的研究によると,実際にネコやサルの視覚野とその周辺で記録される運動感受性細胞は,連続的な運動だけでなく仮現運動にもよく反応する。したがって今日では,知覚は受容器でとらえた感覚信号の空間的・時間的パターンから,中枢神経系で何段階かの情報処理を経て読み取られた,あるまとまった意味のある情報であると理解されている。…
【中脳】より
…上丘はその構造に対応していろいろの機能を行っている。最も重要なものは視覚に関係するものである。上丘は,大脳の後頭葉にある後頭眼野や前頭葉にある前頭眼野などの目の動き(眼球運動)をコントロールしている部分から司令を受けている。…
※「視覚」について言及している用語解説の一部を掲載しています。
出典|株式会社平凡社「世界大百科事典(旧版)」
関連語をあわせて調べる
今日のキーワード
焦土作戦
敵対的買収に対する防衛策のひとつ。買収対象となった企業が、重要な資産や事業部門を手放し、買収者にとっての成果を事前に減じ、魅力を失わせる方法である。侵入してきた外敵に武器や食料を与えないように、事前に...
お知らせ
4/12 日本大百科全書(ニッポニカ)を更新
4/12 デジタル大辞泉を更新
4/12 デジタル大辞泉プラスを更新
3/11 日本大百科全書(ニッポニカ)を更新
2/13 日本大百科全書(ニッポニカ)を更新